If you happen to find yourself in the mountainous regions of Northern Africa or western Arabia, you might catch a glimpse of an Einstein impersonator. However, while the resemblance to Einstein is easy to see, don’t expect theoretical physics from this primate. You might see ancient hieroglyphs portraying this old world monkey as the Egyptian god Thoth, hence this creature’s earned nickname, “sacred baboon” (Contributors, 2014). They are also, interestingly, the only non-human free-ranging primate found in Arabia (Winney et al., 2004).
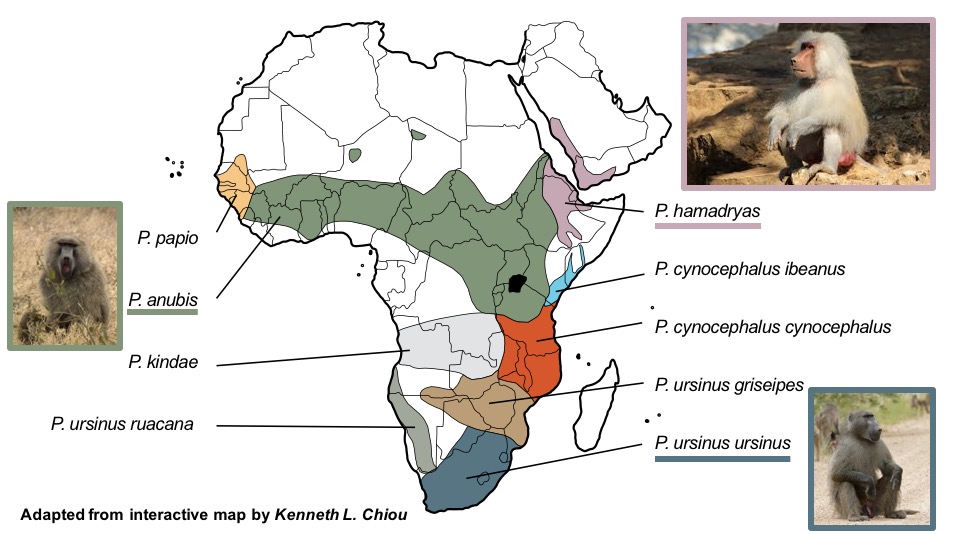
When someone mentions baboons, the hamadryas baboon is probably not the image that comes to mind. Rather, one might picture Rafiki from The Lion King, the cheeky and wise old monkey with a brilliantly-colored face. However, you would be mistaken, as Rafiki is not actually a baboon, but rather a mandrill in the genus Mandrillus and not Papio in which we classify all extant baboon species (Contributors, 2014). These two closely-related taxa are both within the Papionini tribe, and share many morphological features. Understandably, the hamadryas baboon posed difficulties for primatologists when assessing their taxonomic and phylogenetic position in the tribe. Their morphological characteristics are not quite the same of those of a typical savanna baboon. This is largely attributed to their exclusively mountainous habitat, which also has an influence on the distinct socioecological factors contributing to their evolution (D. Zinner et al., 2001).

Like other baboon species, hamadryas baboons are terrestrial, quadrupedal omnivores with buccal cheek pouches (Falk, 2000). These pouches aid the baboons since they largely subsist on low quality vegetation, which is difficult to digest and thus harder to extract necessary nutrients that are abundant in their dry habitats. All baboons have ischial callosities that provide padding when sitting. The ischial callosities are fused in males and unfused in females, allowing for their infamous sexual swellings when in estrus, which occurs non-seasonally in baboons (Contributors, 2014; Falk, 2000). This swelling in the female reproductive organ is an important indicator used by males when determining mating effort. One study suggests that the larger the swelling, the greater the reproductive fitness of the females, and that females with large sexual swellings produce more offspring that survive to reproductive maturity (Domb & Pagel, 2001).

It’s a man’s world for the hamadryas baboons, as their patriarchal hierarchy dictates much of the social life in these large societies. Compared to the savanna baboons populations in which there are much stronger linear female dominance hierarchies within troops, the complex social organization within the hamadryas baboons tend to be male-dominated. A troop of a hamadryas baboons can be over several hundred individuals large. They congregate at the same sleeping site in their mountainous habitat and each of these troops consists of one or more groups who will travel together during the day. The baboons in these smaller groups socialize exclusively among themselves, and these units are known as bands (Schreier & Swedell, 2009). Groups include either solitary males or One Male Units (OMUs), the smallest and most stable social unit. The OMU ranges in size from 2-23 individual baboons, with an average of around 7, with one dominant male that maintains control over several females, his offspring, and on occasion, submissive “follower” males, who are in someway related to the alpha male (Kummer, 1984; Schreier & Swedell, 2009).

While males are typically the dispersing sex in most mammals (i.e. they leave the group they are born in after they reach reproductive age, not unlike humans), in hamadryas baboons, it is the females that disperse from their natal groups and who join a new OMU (Barton et al., 1996). Most of the altruistic behavior seen in hamadryas baboons is directed from females towards males, and they are often observed grooming either the alpha male in their harems, or other males they have befriended. Socioecological theory has conventionally described hamadryas baboons as “non-female bonded” primates, suggesting that females rarely affiliate with each other, since they join new harems after dispersing and thus tend to be unrelated (Hernández-Lloreda et al., 2002). This view is being challenged by recent empirical work that suggests that hamadryas grooming markets might be more complex and influenced by factors such as the number of males in the group and factors that influence activity budgets (Hernández-Lloreda et al., 2002). Thus, there is increased male-male competition within hamadryas baboons, as males have been known to infiltrate neighboring harems at night for a chance to mate with another male’s females (Henzi & Barrett, 2003). If you’ve ever heard the phrase “Hide your wives, hide your kids”, this could be applied to interactions between a male defending his harem from an intruder. These confrontations, more often than not become violent, and infanticide is a common occurrence. Often, the “unfaithful” females are punished by the alpha males who deliver a bite to the back of the female’s neck, often drawing blood and leaving them identifiable due to scars and hair loss around the area (Muller & Wrangham, 2009).

Although all baboon species have extreme sexual dimorphism, the hamadryas baboon takes it to a whole other level, mainly due to the increased male-male intergroup competition. Males are commonly twice the size of females, armed with large canines that are used in displays of dominance (Falk, 2000). Not only are hamadryas baboons sexually dimorphic, but also sexually dichromatic, as males develop silver grey manes around the same time that they reach sexual maturity while smaller females sport contrasting brown hair. Interestingly, if the alpha male drops in rank either due to a harem takeover by a more dominant male or old age, he will lose his silver coat and take on the female’s brown color (Kummer, 2001). Infants, too, differ in fur color as all offspring, regardless of sex, have dark black hair that remains six months after birth, at which point their hair will change to the female color (Shefferly, 2004).

These remarkable primates are not only important to those who reside near their habitats, but also to the scientific community as a whole. Their unique socioecology has fascinated primatologists, as is evident by the long-term research into their social systems and structure, as it is one of most complex among primates. Baboon species, including hamadryas, share many physiological similarities with humans, making these primates vital to studies about human diseases such as epilepsy, heart disease, obesity, and even organ transplantation (Contributors, 2014). Fortunately, according the the IUCN red list, hamadryas baboons are low concern and their populations seem to be increasing. Let’s hope we can continue to conserve this unique baboon species!
Written by Jessica Fernandez, UC Davis undergraduate student athlete in the women’s diving team in her 4th year in the Department of Anthropology. Jessica is a research assistant in Dr. Damien Caillaud’s lab and is interested in topics related to health and disease in primates. She is currently working on a project investigating the gut microbiome of Grauer’s gorillas. This Creature Feature was inspired by a research paper which she wrote for the ANT154A: Primate Socioecology course. After graduating, she hopes to apply to medical school.
Main featured image taken by Hrvoje Sasek [Source]
References:
Barton, R. A., Byrne, R. W., & Whiten, A. (1996). Ecology, feeding competition and social structure in baboons. Behavioral Ecology and Sociobiology, 38(5), 321–329. https://doi.org/10.1007/s002650050248
Contributors, N. W. E. (2014). Baboon- New World Encyclopedia. Retrieved November 26, 2018, from http://www.newworldencyclopedia.org/p/index.php?title=Baboon&oldid=980417
Cowlishaw, G. (1994). Vulnerability To Predation in Baboon Populations. Behaviour, 131(3), 293–304. https://doi.org/10.1163/156853994X00488
Domb, L. G., & Pagel, M. (2001). Sexual swellings advertise female quality in wild baboons. Nature, 410(6825), 204–206. https://doi.org/10.1038/35065597
Falk, D. (2000). Primate Diversity. J. Byram, M. Kelly, M. Babock, & J. Carter (Eds.), W.W. Norton & Company, Inc.
Henzi, P., & Barrett, L. (2003). Evolutionary ecology, sexual conflict, and behavioral differentiation among baboon populations. Evolutionary Anthropology: Issues, News, and Reviews, 12(5), 217–230. https://doi.org/10.1002/evan.10121
Hernández-Lloreda, M. V., Zaragoza, F., & Colmenares, F. (2002). Grooming and coercion in one-male units of hamadryas baboons: market forces or relationship constraints?. Behaviour, 139(11-12), 1525-1553.
Kamilar, J. M. (2006). Geographic Variation in Savanna Baboon (Papio) Ecology and its Taxonomic and Evolutionary Implications. In Primate Biogeography (pp. 169–200). Springer US. https://doi.org/10.1007/0-387-31710-4_6
Kummer, H. (1984). From laboratory to desert and back: A social system of hamadryas baboons. Animal Behaviour, 32(4), 965–971. https://doi.org/10.1016/S0003-3472(84)80208-0
Kummer, H. (2001) A Male Dominated Society: The Hamadryas Baboon of Cone Rock, Ethiopia. In The Encyclopedia of Mammals (pp. 376-77), 2nd edition. MacDonald, D. (Ed.) Oxford University Press.
Muller, M. N., & Wrangham, R. W. (2009). Sexual coercion in primates and humans : an evolutionary perspective on male aggression against females. Harvard University Press.
Newman, T. K., Jolly, C. J., & Rogers, J. (2004). Mitochondrial phylogeny and systematics of baboons (Papio). American Journal of Physical Anthropology, 124(1), 17–27. https://doi.org/10.1002/ajpa.10340
Shefferly, N. 2004. “Papio hamadryas” (On-line), Animal Diversity Web. Accessed February 15, 2019 at https://animaldiversity.org/accounts/Papio_hamadryas/
Schreier, A. L., & Swedell, L. (2009). The fourth level of social structure in a multi-level society: ecological and social functions of clans in hamadryas baboons. American Journal of Primatology, 71(11), 948–955. https://doi.org/10.1002/ajp.20736
The IUCN Red List of Threatened Species. (n.d.). Retrieved from https://www.iucnredlist.org/
Winny, B. J., Hammond, R. L., Macasero, W., Flores, B., Boug, A., biquand, V., … Bruford, M. W. (2004). Crossing the Red Sea: phylogeography of the hamadryas baboons, Papio hamadryas hamadryas. Molecular Ecology, 13(9), 2819–2827. https://doi.org/10.1111/j.1365-294X.2004.02288.x
Zinner, D., Peláez, F., & Torkler, F. (2001). Distribution and Habitat Associations of Baboons (Papio hamadryas) in Central Eritrea. International Journal of Primatology, 22(3), 397–413. https://doi.org/10.1023/A:1010703611820
Zinner, D., Wertheimer, J., Liedigk, R., Groeneveld, L. F., & Roos, C. (2013). Baboon phylogeny as inferred from complete mitochondrial genomes. American Journal of Physical Anthropology, 150(1), 133–140. https://doi.org/10.1002/ajpa.22185

{kind=link}
{kind=link}
{kind=link}
{kind=link}