During the summer solstice, animals that inhabit regions north of the Arctic Circle can experience weeks to months of continuous polar illumination. For migratory birds breeding at higher latitudes, this extreme photoperiodicity—in conjunction with intense competition for mates and territory—can lead to decrements in sleep. While animals deprived of sleep typically show reduced neurobiological performance, health, and in some cases, survival, several vertebrate taxa exhibit adaptations that permit normal activities even when sleep is reduced. However, little work has been done to determine whether behavioral costs of sleep loss—commonly seen in other organisms, such as humans—occur in arctic-breeding songbirds.
In 2019, two researchers at Western Kentucky University (WKU) aimed to better understand the behavioral costs of reduced sleep by investigating the effect of sleep loss on cognitive performance in an arctic-breeding songbird, the Lapland longspur (Calcarius lapponicus)—a species that experiences 24 hours of constant polar illumination and exhibits around-the-clock activity during its short breeding season, with an inactive period of approximately four hours per day.

To test whether longspurs exhibit behavioral costs of sleep loss, researchers trained individually housed birds to complete two executive function tasks; a color association task and a spatial learning task [1]. For the color association task (Fig. 1a), a food reward (mealworm; Tenebrio molitor) was randomly placed in one of the wells of a 6-well cell culture plate and covered by a plastic blue chip. The criterion for success was for birds to uncover the mealworm by removing the blue chip from atop the rewarded well within 10 minutes before touching the white chip, which covered an unrewarded well. For the spatial learning task (Fig. 1b), a mealworm was placed in the back left well and covered by a plastic red chip. A red cue card (Fig. 1b; yellow arrow) was placed behind the rewarded well to orient birds within their cages. The criterion for success was for birds to uncover the mealworm by removing the red chip from atop the rewarded well within 10 minutes without touching the other three red chips, which covered unrewarded wells.

Once birds successfully completed both the color association and spatial learning tasks every day for two weeks, individuals were considered to have effectively committed each task to memory. Next, birds were deprived of sleep for 24 hours using an automated sleep fragmentation machine (see Hodinka & Ashley, 2020) and administered both tasks a final time to measure the effect of sleep loss on memory recall. During the final test, birds were video recorded and scored based on task completion (correct/incorrect) and time to completion. Notably, to ensure birds were not using sensory cues (e.g., hearing, smell) to find the food reward, mealworms were removed on the final test day.
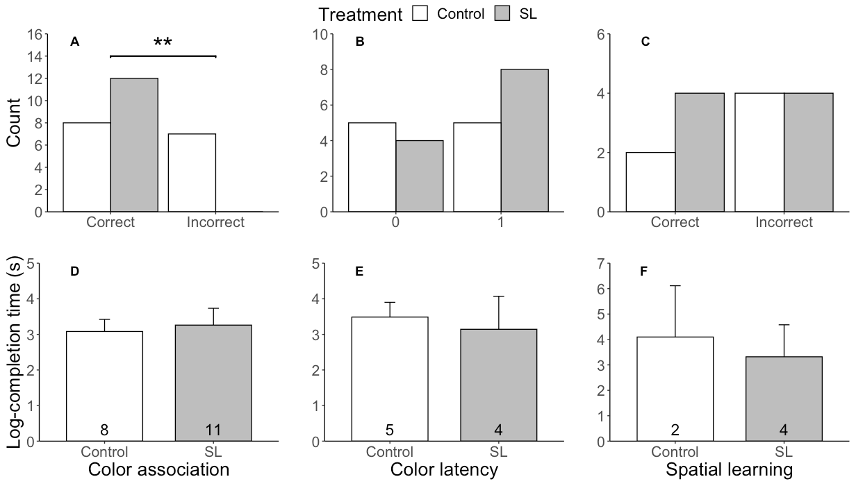
Researchers found birds that had undergone the 24-hour sleep loss (hereafter SL) protocol performed significantly better on the color association task relative to controls (Fig. 2A; P < 0.01). All 12 SL birds completed the task correctly while only half of the control birds completed the task correctly. However, there was no significant difference in time to complete the color association task (Fig. 2D). Notably, a secondary task linked to the color association task termed “color latency” was quantified during the final test, post-SL. This was not a task that birds were explicitly taught; instead, during the color association task, researchers quantified (1) whether birds attempted to flip over the white chip after first removing the blue chip and (2) time taken to remove the white chip. The goal was to investigate the degree of plasticity in a food-driven memory task and whether SL played a role in an individual’s choice of foraging elsewhere. In this case, there was no significant difference in the choice made on the color latency task (Fig. 2B; 0 = white chip removed, 1 = white chip not removed) nor time taken to remove the white chip after first removing the blue chip (Fig. 2E). Additionally, for the spatial learning task, researchers found no significant difference in the success rate (Fig. 2C) nor time taken to remove the back left chip (Fig. 2F).
Not only do the results obtained from the color association task align with current literature, but there may be an adaptive role for corticosterone in ameliorating the cognitive deficits of SL as these birds had significantly elevated corticosterone concentrations (P < 0.05) relative to controls. However, results from the spatial learning task appear to be more enigmatic. While it is well known that, in humans, non-spatial memory is stored in the prefrontal cortex, spatial memory is stored in the hippocampus which is thought to be more sensitive to the effects of SL. That said, if longspurs are behaviorally insensitive to the effects of SL, it seems paradoxical that decrements in spatial memory would be observed compared to nonspatial memory. During the breeding season, longspurs presumably rely upon spatial memory to navigate the tundra landscape with limited markers for spatial orientation. However, it is possible they use other nonspatial cues to successfully locate territories and nest sites. In the future, researchers aim to assess whether temperate-breeding birds that breed in regions precluding 24 hours of sunlight, such as the chestnut-collared longspur (Calcarius ornatus), are more sensitive to the behavioral and physiological effects of sleep loss relative to polar-breeding birds.
For more information:
Lapland longspurs completing the color association task
Hodinka, B.L., & Ashley, N.T. (2020). Effect of sleep loss on executive function and plasma corticosterone levels in an arctic-breeding songbird, the Lapland longspur (Calcarius lapponicus). Hormones and Behavior, 122:104764.
References:
[1] Anderson, R.C., Searcy, W.A., Peters, S., Hughes, M., DuBois, A.L., & Nowicki, S. (2017). Song learning and cognitive ability are not consistently related in a songbird. Animal Cognition, 20, 309–320.
[All images provided by Brett Hodinka; Edited by Lindsey Broadus]
